GLI ACIDI GRASSI DEL LIEVITO: STRUTTURA E CARATTERISTICHE
Il lievito enologico per eccellenza, Saccharomyces cerevisiae, come tutti gli organismi unicellulari, delimita il citoplasma dall’ambiente esterno tramite una membrana cellulare; anche il nucleo della cellula è delimitato da una membrana nucleare. La membrana cellulare è posta all’interno della parete cellulare, con cui non va confusa. La parete cellulare è composta da polisaccaridi, tra cui le mannoproteine, ed ha uno scopo di delimitazione con il mezzo in cui il lievito sviluppa, ma è uno scopo principalmente meccanico. La membrana cellulare, invece, ha una funzione fisiologica di primario rilievo. Specialmente la sua integrità è di fondamentale importanza nelle fasi finali della fermentazione alcolica, in quanto l’alcol ha la capacità di assottigliarla fino a farle perdere il 30% del suo spessore.
La membrana cellulare è polare al suo esterno, mentre è apolare al suo interno. I mattoni della membrana cellulare sono i fosfolipidi, molecole complesse che possono essere schematizzate nella seguente immagine.

Nella figura in alto, seppure in modo semplificato, si osserva la componente fosfolipidica della membrana cellulare. In verde sono rappresentati gli acidi grassi; in giallo, la componente non lipidica.
L’acido grasso più lungo, con 18 atomi di carbonio è l’acido stearico, che è un C18:0, in quanto ha 18 atomi di carbonio e 0 insaturazioni. L’assenza di insaturazioni, ovvero di doppi legami tra carbonio e carbonio determina la linearità della molecola.

Un altro acido grasso di rilevante importanza è l’acido palmitico, che è un C16:0, avendo 16 atomi di carbonio e nessuna insaturazione. Come si può vedere dalle formule di struttura gli acidi grassi saturi, ovvero con zero insaturazioni, sono lineari. Gli acidi grassi saturi sono presenti nei derivati animali (es il burro). Gli acidi grassi insaturi sono presenti nei derivati vegetali (olio d’oliva, olio di lino). La regola prevede, come sempre accade in natura, delle eccezioni.

Oltre agli acidi grassi saturi, ovvero lineari, vi sono gli acidi grassi monoinsaturi, che presentano una curva sul punto di insaturazione (ovvero il doppio legame). L’acido oleico ha 18 atomi di carbonio ed una insaturazione sul C9. L’acido palmitoleico ha 16 atomi di carbonio ed una insaturazione sul carbonio 9 (C9).

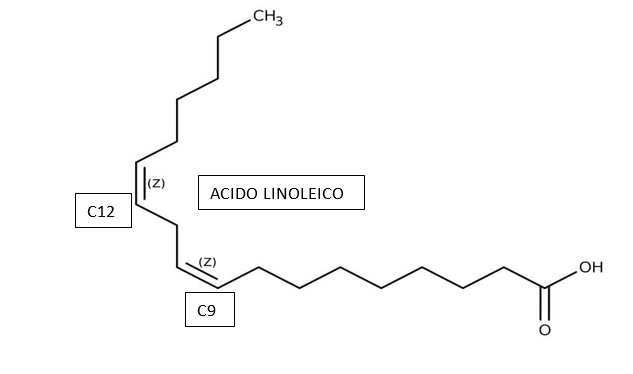
Esiste poi una piccola frazione di acidi grassi a doppia insaturazione, ovvero l’acido linoleico che è un C18:2 con insaturazione su C9 e C12. L’acido linoleico è noto per essere un omega – 6.
Nella membrana cellulare vi sono diversi acidi grassi e un sunto, per quanto indicativo, è riportato nella tabella sottostante. Notiamo che vi sono piccole percentuali di acidi grassi a media catena, di cui parleremo dopo. Il più rappresentato pare essere l’acido palmitico C16:0, seguito dalla forma cis dell’acido oleico C18:1; vi è una piccola percentuale di acido linoleico C18:2.

Come si formano gli acidi grassi nella cellula del lievito? Il precursore è il glucosio, da cui, via glicolisi si forma l’acido piruvico, e per decarbossilazione il radicale acilico che viene legato al coenzima A. Il coenzima A è un trasportatore di radicali acilici. Il radicale acilico è un acido carbossilico privato dell’ossidrile

Dall’unione tra gruppo acetile e il coenzima A si ottiene l’acetil coenzima A. Nell’immagine sottostante è raffigurata la molecola, con il gruppo acetile evidenziato in rosso.

Tramite l’enzima FAT (FATTY ACID SYNTHASE) i mattoni degli acidi grassi vengono saldati tra loro a formare delle catene di 16 o 18 atomi di carbonio, tramite l’azione del coenzima A. Al fine di ottenere acidi grassi a lunga catena (C16, C18) è necessario che il lievito possa innanzitutto spostare i radicali acetilici tramite il coenzima A: è quindi necessario che nel mosto ci sia fosforo, azoto e zolfo, che oltre al carbonio sono i mattoni fondamentali della molecola. Il successivo step nella formazione degli acidi grassi è la carbossilazione dell’acetil coenzima A, da cui si ottiene il malonil coenzima a A: nella figura sottostante l’acile dell’acido malonico è evidenziato in verde.
Cercando di semplificare la reazione possiamo dire che la componente polare selezionata in verde viene addizionata di volta in volta di due etili (previa eliminazione degli ossigeni) che vengono trasportati dal coenzima A sotto la forma di acetili (selezionati in rosso).
Come avviene pure nel corpo umano è con questa via metabolica che una parte del glucosio viene trasformata in acidi grassi e negli animali in trigliceridi che vanno a costituire il tessuto adiposo.
È stato dimostrato che un mezzo ricco in acidi grassi di origine vegetale, (nel nostro caso le bucce dell’uva) principalmente linoleico e linolenico, rende più funzionale le membrana del lievito, migliorandone il metabolismo. In poche parole, un mosto più verde e più torbido, aiuta il lievito a fermentare.
A parità di altri fattori, quali zuccheri, azoto disponibile, pH la fermentazione ad alte NTU permette al lievito migliori performance. Questo fatto è facilmente osservabile in cantina: i mosti con NTU inferiori a 20 peggiorano la funzionalità di membrana del lievito: il lievito utilizza meno l’azoto alfa amminico e tende a produrre più acido acetico, infine, sono più probabili gli arresti fermentativi.
Uno dei motivi è il fatto che a basse torbidità il lievito produce un maggiore tenore di acidi grassi a media catena, acido decanoico e ottanoico, che sono dei veri e propri inibitori della fermentazione. Ammetto che in letteratura enologica, pur essendo in vari articoli citato il fatto che l’acido decanoico è dannoso per il lievito, non vi è molta chiarezza sulla via metabolica che porta alla formazione di questo acido.
Ma, essendo rilevato il fatto che esso viene formato maggiormente in mezzi molto limpidi, è ipotizzabile che il lievito in tali condizioni abbia minori mattoni per costruire acidi grassi a lunga catena (C16, C18), e si fermi quindi ad acidi a soli 6, 8 o 10 atomi di carbonio. Gli acidi grassi prodotti dal lievito hanno sempre un numero pari di atomi di carbonio, proprio perché il lievito tramite il Coenzima A aggiunge degli acetili, ovvero 2 carboni alla volta.
Gli acidi grassi a corta catena sono:
C2, acido acetico o etanoico
C4, acido butirrico o butanoico
C6, acido esanoico
Gli acidi a media catena sono:
C8, acido ottanoico
C10, acido decanoico
C12, acido dodecanoico.
In conclusione l’obbiettivo enologico deve essere quello di favorire la produzione di acidi grassi a lunga catena, con una discreta percentuale di acidi grassi insaturi, che migliorano il funzionamento della membrana cellulare.
E’ auspicabile che ulteriori studi vengano svolti in questo campo.